A Binding Advantage: Optibumin 25 Demonstrates Enhanced Drug Binding and Consistency over Plasma Albumin
Published on 15 October 2025
Application Note
EXECUTIVE SUMMARY
Human serum albumin (HSA) is exemplary among proteins in drug complexation applications. It is capable of binding a wide variety of ligands with high affinity, is resistant to renal filtration, has a lengthy in vivo half-life, is incredibly soluble, and contains multiple drug binding sites. These unique pharmacological properties of albumin are due to its structure, but a protein’s structure is complex and heavily influenced by its environment. Because clinical HSA is donor-derived and stabilized with additives, it is heterogeneous and contains aggregates making it suboptimal for high precision applications like drug complexation. To address these shortcomings, InVitria has developed Optibumin 25, a chemically defined, animal-origin-free, excipient-grade, cGMP, recombinant human serum albumin (rHSA). In this application note, we demonstrate that Optibumin 25 has higher binding affinity than HSA for a model drug demonstrating its superiority for precise drug complexation applications.
Key Highlights
- Nearly 3x higher drug-binding affinity vs. plasma HSA
- 10x fewer aggregates (p < 0.05)
- 99% reduced Cys34 vs. ~60% in plasma HSA
- No stabilizers that interfere with binding
- Consistent lot-to-lot performance

INTRODUCTION
Albumin is unique among proteins in its ability to bind a plethora of ligands. Classic examples of albumin binding drugs include small molecules such as ibuprofen (Evoli et al., 2017), caffeine (Banipal et al., 2016), and warfarin (O’Reilly, 1967). More recent and innovative examples include the chemotherapeutics Abraxane, [(Desai et al., 2006), (Miele et al 2009), (Green et al 2006)] and Fyarro (Zhou et al 2025), the glucagon like peptide 1 (GLP-1) agonist Semaglutide (Lau et al., 2018) and lipid conjugate siRNAs [(Hoogenboezem et al., 2024), (Sarett et al., 2017), (Colazo et al., 2025)]. Albumin is also a major transporter of important natural bioactive molecules such as lipids, steroid hormones, metal ions and heme (Mishra et al., 2021), (Ashraf et al 2023). Albumin binding to these ligands endows them with many pharmacological benefits.
Complexing with albumin improves drug bioavailability and reduces the need for repeated dosing by improving the in vivo half-life of complexed drugs. There are two main reasons for this benefit. First, with a molecular weight of 66 kDa, albumin and its complex partners are resistant to renal filtration which rapidly clears free molecules smaller than 30-50 kDa (Ruggiero et al., 2010) including peptides, small molecules, and siRNAs. Second, albumin contains a domain that binds the neonatal Fc receptor (FcRn)enabling recycling by the same mechanism that allows long term preservation of IgG in serum (Hoogenboezem and Duvall 2018). This recycling gives both albumin and IgG a remarkable 19-day serum half-life (Liu et al., 2020).
Albumin binding also improves the solubility and reduces the toxicity of its complex partners (Spada et al., 2021). Fatty acids and small molecule drugs are generally hydrophobic and poorly water soluble. Therefore, they are often solubilized in organic solvents like ethanol, DMSO, or DMF but these solvents pose safety risks in vivo (Madsen et al., 2019). Fortunately, albumin has a high binding capacity for these molecules including two main sites for ligand binding termed Sudlow sites I and II (Sudlow et al., 1975) as well as a more generalized site termed simply drug binding site III (Kraugh-Hansen et al., 1983) in addition to seven fatty acid binding sites (Fujiwara et al., 2007). This high binding capacity for ligands obviates the need for toxic solvents.
This amount of hydrophobic cargo would cause many proteins to precipitate, but unlike most proteins, albumin is exceptionally soluble and stable thanks to a globular structure containing 17 disulfide bonds and a strong negative charge at physiological pH (He and Carter, 1992). Albumin’s exceptional solubility also enables it to reach very high concentrations in plasma. This allows a substantial reservoir of albumin-bound drug to persist in the plasma and dissociate at a slow controlled rate over time to maintain active free drug concentration over time (Larsen et al., 2016).
Reaping the benefits of albumin drug complexation relies on strong and consistent drug binding which depends on albumin’s structure. Unfortunately, serum albumin’s structure is intricate and can be influenced by many factors. For example, post-translational modifications (PTMs) such as cysteinylation and glycation are affected by donor age and blood sugar levels respectively (Era et al., 1995), (Rondeau and Bourdon 2011) which can affect drug binding (Lee and Wu, 2015), (Nagumo et al., 2014) as well as FcRn binding (Leblanc et al., 2019), (Ward et al., 2022). Formulation can also impact albumin structure. For instance, the stabilizers sodium octanoate (also known as caprylate or octanoic acid) and N-acetyl tryptophan are added to plasma albumin to stabilize the protein during pasteurization which is required to inactivate bloodborne pathogens. These stabilizers bind to Sudlow site II (Faroongsarng and Kongprasertkit, 2014) which has been shown to interfere with drug binding to the same site (Tramarin et al., 2019), (Kragh-Hansen 1991), (Harm et al., 2018), (Olsen et al., 2004). Aggregation may also impact binding by reducing the surface area available for complexation. All of these factors contribute to pharmacokinetic variation in drug formulation.
To address these issues, InVitria has developed Optibumin 25, a recombinant, animal-free albumin that is free of stabilizers and aggregates and has few post-translational modifications. Here we compare the binding affinity of Optibumin and plasma HSA for warfarin, a model prescription blood thinning drug that binds to Sudlow site I (Petitpas et al., 2001).
RESULTS AND DISCUSSION
Optibumin 25 has Superior Drug Binding Capacity Compared to HSA
To quantitate the binding affinity of warfarin to Optibumin 25 or HSA, various concentrations of the drug were mixed with constant amounts of either albumin and UV-Vis spectra were recorded to measure total warfarin. Next, free and bound warfarin were separated by low MWCO spin filtration and the retentates and filtrates characterized by UV-Vis to measure bound and free drug respectively.
This data was transformed to create a Scatchard plot (Figure 1). Briefly, the Scatchard model assumes that a ligand (e.g. warfarin) and a target (e.g. albumin) each with one binding site interact reversibly to form a complex with a dissociation constant KD. The plot is a linearized transformation of this relationship where the slope is -1/KD meaning that a steeper slope is indicative of stronger binding (Scatchard, 1949). Construction of this plot is explained in more detail in the materials and methods.
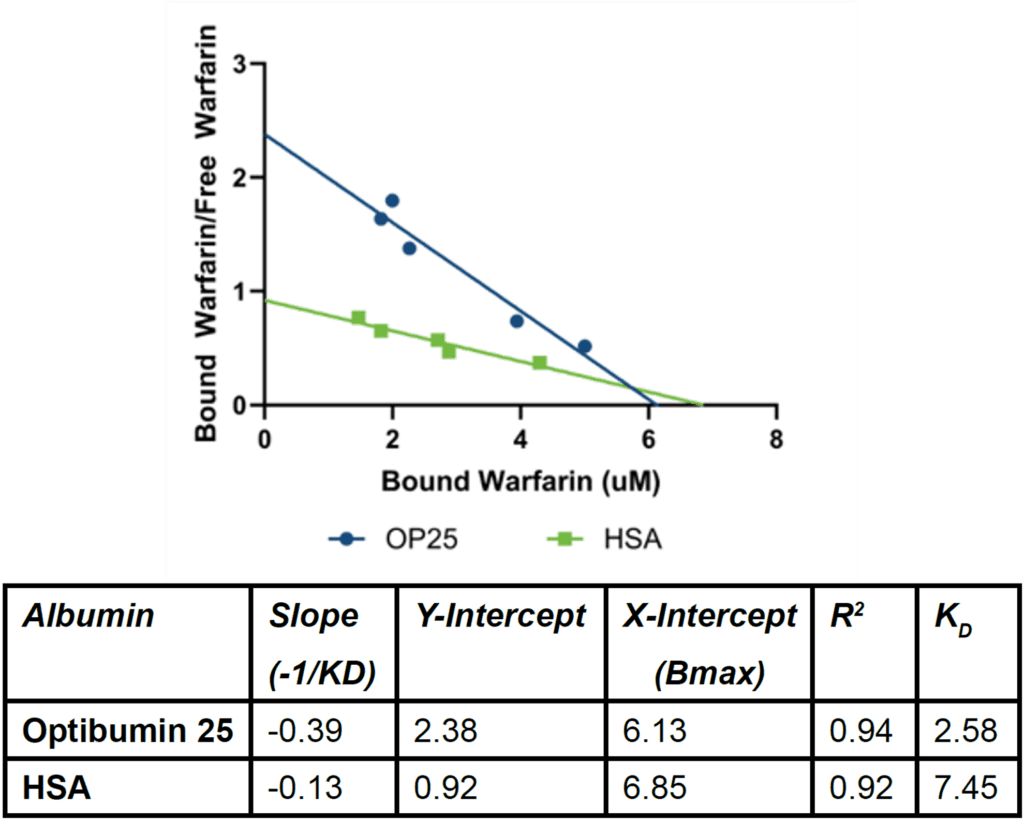
Because the slope of the Optibumin plot is significantly steeper than that of the HSA plot (Fisher’s F-test p < 0.01), Optibumin binds warfarin more potently. The measured KD value of 2.575 µM for Optibumin aligns with the expected 3 µM affinity reported in the literature (Sudlow et al., 1975), (Diana et al., 1989) (Loun and Hage, 1994) whereas the KD for serum HSA and warfarin, 7.45 µM, was significantly higher indicating that Optibumin binds warfarin exactly as well as predicted whereas HSA exhibits lower than expected binding. Additionally, because the x-
intercepts for both regressions are similar, Optibumin and HSA have similar total binding capacities for warfarin.
Importantly, more potent binding would translate to a more significant reservoir effect and a longer in vivo half-life for warfarin in complex with Optibumin versus HSA (Fan et al., 2022). This suggests that Optibumin would retain warfarin for a longer time than serum albumin, causing a more gradual, controlled release of active free drug over time. This means that doses could potentially be raised with less risk of toxicity, or doses could be given less frequently (Dennis et al., 2002). For drugs with a narrow therapeutic index (the ratio of a drug’s toxic concentration divided by its effective concentration) such as warfarin (Johnson, 2012) the clinical implications of this could be substantial.
Because warfarin and the stabilizers octanoate and N-acetyl tryptophan all target defined binding sites on albumin, it is expected that they would compete either directly or allosterically for these sites (Yamasaki et al., 1999), (Ghuman et al., 2005). This means that Optibumin, which is uniquely free of these stabilizers (Table 1) likely binds to a wide variety of albumin complexing drugs with higher affinity than HSA in addition to warfarin.

Optibumin 25 Has Fewer Aggregates and Post Translational Modifications Than HSA
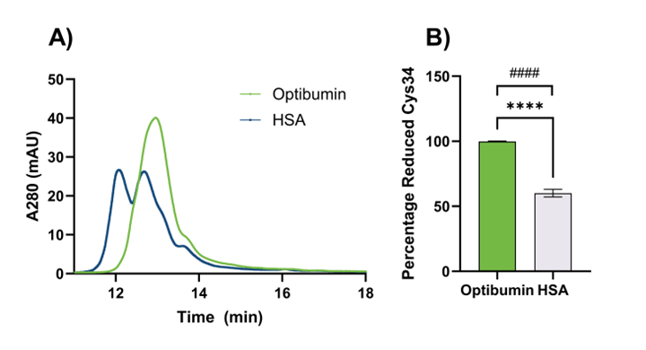
Pinpointing the precise mechanism for Optibumin’s improved binding affinity is beyond the scope of this work but we offer several hypotheses – which are not mutually exclusive – that could potentially explain this phenomenon. First, it could be that differences in the amount of aggregated albumin present in Optibumin and plasma HSA contribute to this because aggregated albumin has less surface area available to interact with free drug molecules. This hypothesis is supported by our previous observations of differences in aggregation of Optibumin and HSA by capillary electrophoresis (Figure 2A). The extent of aggregation from the gel image is quantified in Figure 2B. In a sample of 3 lots of Optibumin and 5 lots of HSA, we found that Optibumin had a mean of 39000 area units per µg of protein compared to 403000 area units per µg of protein for HSA, an astounding 10-fold difference that was statistically significant (Welch’s t test p < 0.05). Furthermore, Optibumin had a standard deviation of 11000 area units per µg of protein while HSA had a standard deviation of 257000 area units per µg of protein which was also a significant difference (F test for variance, p < 0.01). This demonstrates that Optibumin is more consistent than clinical HSA which is paramount when trying to adjust dosing regimens of drugs with a narrow therapeutic window to maintain efficacy and avoid toxicity (Celestin et al., 2022).

Second, it could be that bound stabilizers or post translational modifications on plasma albumin interfere with the drug interaction. The presence of stabilizers has been directly linked to reduced ligand binding capacity (Park et al., 2021). As an example of the effect of posttranslational modifications on binding, cysteinylation of albumin’s lone free cysteine, Cys34, has been implicated in reducing warfarin binding (Nagumo et al., 2014). We have previously shown that plasma HSA is 40% cysteinylated whereas Optibumin is 99% unmodified at this residue (Figure 3) and observed significant differences in both mean and variance of cysteinylation between Optibumin and plasma HSA. This superior consistency can help improve predictability in the clinic which could lead to better outcomes (Fender et al., 2019). Fatty acid binding or other post translational modifications such as glycation can also contribute to this effect (Vorum et al., 1993). Furthermore, in some cases ligands that bind to one Sudlow site such as octanoate, can allosterically affect binding to the other Sudlow site (Yamasaki et al., 1999), (Ghuman et al., 2005).
CONCLUSION
InVitria’s Optibumin 25 recombinant albumin was shown to have a higher affinity than HSA for the model albumin binding drug warfarin. This improved binding could have important pharmacological benefits such as enabling lower or less frequent drug dosing or improving safety and efficacy. It is likely this improved binding is due to differences in: formulation (i.e. the presence of stabilizers in HSA but not Optibumin), post-translational modifications (the presence of cysteinylated albumin in HSA but not Optibumin), and/or aggregation which was significantly lower in Optibumin than HSA. Furthermore, Optibumin also demonstrated more consistency in aggregation and oxidation state, suggesting that complexing drug to Optibumin could improve pharmacokinetics, which could have clinical benefits. Taken together, these factors along with its recombinant animal-origin-free nature, make Optibumin 25 the superior albumin for drug complexation applications.
MATERIALS & METHODS
Warfarin Standard Curve Development
Warfarin sodium salt was prepared at a stock concentration of 1 mg/mL in deionized water. The stock was then serially diluted 2-fold to get concentrations spanning 4 orders of magnitude. The UV-Vis absorbance spectrum was recorded for each sample to generate a standard curve and define the linear range of the absorbance at the spectral peak of 310 nm to enable quantitation of free warfarin.
Albumin Preparation
Plasma HSA was desalted by spin filtration to remove N-acetyl tryptophan which interferes with absorbance measurements at 280 nm. This was not necessary for Optibumin because it contains no such additives. Stocks of Optibumin and HSA were prepared at 0.2 mg/ml as measured by A280 were prepared for complexing reactions.
Albumin-Drug Complexing
To evaluate the binding affinity of albumin to warfarin, Optibumin or HSA were first mixed 1:1 with serial dilutions of warfarin. These dilutions spanned the pre-established linear range of warfarin concentrations detectable by UV-Vis for a final albumin concentration of 0.1 mg/mL and a final sample volume of 1 mL. The absorbance spectra of the samples were then recorded. This represented the total warfarin (free and bound together). The samples were then buffer exchanged by spin filtration through a 3 kDa MWCO membrane. The flowthrough and retentate were each adjusted to total volumes of 1 mL each and quantified by UV-Vis. The flowthrough comprised free warfarin and the retentate comprised bound warfarin. The contribution of albumin to the total and bound warfarin was subtracted.
Scatchard Plot Construction and Interpretation
The strength of binding of a drug to a target is defined as KD = koff/kon where koff and kon are the kinetic rate constants governing dissociation and association of the drug to the target respectively. The lower the KD, the less dissociation and the stronger the binding affinity. Drug binding can be plotted as the proportion of total drug that is bound, on the y-axis vs free drug on the x-axis to get an isothermal binding curve (Weber et al., 2020). On these plots, KD is the point on the x-axis where the y value is half its maximum. However, because these plots are nonlinear, interpretation can be challenging, and accurate fitting requires many data points. The Scatchard plot addresses these issues by linearizing the relationship between the drug and target concentrations. This transformation is relatively straightforward and given below.
The Scatchard model relies on reversible association and dissociation of a free drug D and unbound target T to form a drug target complex DT.

According to the law of mass action, the rate of reaction depends on the kinetic rate constants and the concentrations of reactants. If we assume the reaction is at equilibrium i.e. association and dissociation occur at the same rate, then we have:
![Binding equilibrium condition showing kon[D][T] equals koff[DT].](https://invitria.com/wp-content/uploads/2026/03/Equasion-2-300x65.png)
Using the definition of KD we have:
![Definition of equilibrium dissociation constant KD as koff divided by kon and equal to concentration ratio [D][T]/[DT].](https://invitria.com/wp-content/uploads/2026/03/Equasion-3-300x92.png)
Let nbar be the average number of sites on the target bound by the drug and let Ttotal be the total number of target sites
![Fractional occupancy equation defining average bound fraction n̄ as [DT] divided by total target concentration.](https://invitria.com/wp-content/uploads/2026/03/Equasion-4-300x84.png)
Solving for the concentration of free target sites:
![]()
Rearranging the equation for KD by substituting nbar

Rearranging further to solve for nbar /D:
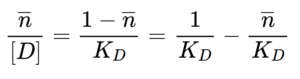
This is the Scatchard equation in which nbar/D, the ratio of bound to free drug is on the y-axis, nbar the proportion of drug that is bound, is on the x-axis, -1/KD is the y-intercept and -n/KD is the slope. While the axes may not be immediately intuitive, the advantage is that KD, the most critical parameter for drug binding, can easily be determined from the slope of the plot.
Aggregate Analysis
Liquid samples of Optibumin 25 and HSA were diluted down to 1 mg/mL and 0.5 mg/mL using 20 mM phosphate buffer, 50mM NaCl, pH 7. For the powdered sources, HSA source 3 and source 4, samples were reconstituted initially at 5-10 mg/mL and diluted down to 1 and 0.5 mg/mL using 20 mM phosphate buffer, 50 mM NaCl, pH 7. These diluted samples were subsequently used for quantifying albumin aggregate using chemiluminescence-based capillary electrophoresis on a Wes system (ProteinSimple, San Jose, CA). Samples were prepared under native conditions by reconstituting the fluorescent master mix supplied by ProteinSimple in water only (no DTT or SDS sample buffer were added) and mixing with the albumin samples at a 1 to 5 ratio (1 part fluorescent master mix and 5 parts diluted albumin samples). Albumin aggregate was analyzed using the Compass for SW software (version 6.3.0). The aggregate area (any detection above the main monomer band at ~60 kDa) was quantified using dropped line peak analysis. For each sample, the total aggregate area was divided by the diluted concentration, either 1 mg/mL or 0.5 mg/mL, to obtain the “aggregate band area per injected mass.” Statistical analysis was run to show that aggregate area was significantly lower for Optibumin (Welch’s t test) and more consistent lot to lot than HSA (f test for equality of variance).
Applications & Use Cases
Optibumin 25 enhances formulation performance and reproducibility across a wide range of development and manufacturing applications:
- Biologics formulation – improves drug stability and binding uniformity
- Small-molecule complexation – increases solubility and control of release kinetics
- Cell and gene therapy manufacturing – provides consistent, regulatory-compliant excipient performance
- Vaccine and therapeutic protein formulation – minimizes variability from donor-derived materials
Regulatory Relevance
- Animal-origin-free (AOF): eliminates plasma-derived variability and risk
- cGMP-manufactured: designed for clinical and commercial use
- Regulatory alignment: meets FDA and EMA guidance for eliminating donor-derived raw materials in biologics
Featured Solution
Optibumin 25 – Recombinant Human Serum Albumin
Optibumin 25 is a high-purity recombinant albumin designed for drug formulation, biologics manufacturing, and pharmaceutical development. With its strong binding performance, minimal aggregation, and lot-to-lot consistency, Optibumin 25 provides a superior foundation for safer, more predictable therapies.
Frequently Asked Questions (FAQs)
Q: What makes Optibumin 25 different from plasma-derived HSA?
Optibumin 25 is a recombinant human serum albumin (rHSA) produced under animal-origin-free, cGMP conditions. Unlike plasma-derived HSA, it contains no stabilizers or donor-related variability, resulting in greater purity, lower aggregation, and more consistent drug-binding performance.
Q: How does improved drug binding impact pharmaceutical formulations?
Higher binding affinity supports better solubility, stability, and pharmacokinetic control. Optibumin 25’s strong and predictable binding enables more reliable drug complexation and can reduce variability in formulation development and therapeutic performance.
Q: What types of applications can benefit from Optibumin 25?
Optibumin 25 is ideal for drug formulation, biologics, vaccines, and gene or cell therapy manufacturing. Its purity and consistency make it a reliable excipient for both clinical development and commercial manufacturing.

See the full dataset comparing recombinant and plasma-derived albumin in drug complexation.
The following content is gated. Please, subscribe to open access to it.
Footnotes
- Ashraf, S., Qaiser, H., Tariq, S., Khalid, A., Makeen, H. A., Alhazmi, H. A., & Ul-Haq, Z. (2023). Unraveling the versatility of human serum albumin: A comprehensive review of its biological significance and therapeutic potential. Current Research in Structural Biology, 6, 100114. https://doi.org/10.1016/j.crstbi.2023.100114
- Banipal, T. S., Kaur, N., & Banipal, P. K. (2016). Binding studies of caffeine and theophylline to bovine serum albumin: Calorimetric and spectroscopic approach. Journal of Molecular Liquids, 223, 1048–1055. https://doi.org/10.1016/j.molliq.2016.09.034
- Celestin, M. N., & Musteata, F. M. (2021). Impact of changes in free concentrations and drug–protein binding on drug dosing regimens in special populations and disease states. Journal of Pharmaceutical Sciences, 110(10), 3331–3344. https://doi.org/10.1016/j.xphs.2021.05.018
- Colazo, J. M., Keech, M. C., Shah, V., Hoogenboezem, E. N., Lo, J. H., Francini, N., Cassidy, N. T., Yu, F., Sorets, A. G., McCune, J. T., DeJulius, C. R., Cho, H., Michell, D. L., Maerz, T., Vickers, K. C., Gibson-Corley, K. N., Hasty, K. A., Crofford, L. J., Cook, R. S., & Duvall, C. L. (2025). siRNA conjugate with high albumin affinity and degradation resistance for delivery and treatment of arthritis in mice and guinea pigs. Nature Biomedical Engineering, 9, 1366–1383. https://doi.org/10.1038/s41551-025-00945-1
- Dennis, M. S., Zhang, M., Meng, Y. G., Kadkhodayan, M., Kirchhofer, D., Combs, D., & Damico, L. A. (2002). Albumin binding as a general strategy for improving the pharmacokinetics of proteins. Journal of Biological Chemistry, 277(38), 35035–35043. https://doi.org/10.1074/jbc.M205854200
- Desai, N., Trieu, V., Yao, Z., Louie, L., Ci, S., Yang, A., Tao, C., De, T., Beals, B., Dykes, D., Noker, P., Yao, R., Labao, E., Hawkins, M., & Soon-Shiong, P. (2006). Increased antitumor activity, intratumor paclitaxel concentrations, and endothelial cell transport of cremophor-free, albumin-bound paclitaxel (ABI-007), compared with cremophor-based paclitaxel. Clinical Cancer Research, 12(4), 1317–1324. https://doi.org/10.1158/1078-0432.CCR-05-1634
- Diana, F. J., Veronich, K., & Kapoor, A. L. (1989). Binding of nonsteroidal anti-inflammatory agents and their effect on binding of racemic warfarin and its enantiomers to human serum albumin. Journal of Pharmaceutical Sciences, 78(3), 195–199. https://doi.org/10.1002/jps.2600780304
- Evoli, S., Mobley, D. L., Guzzi, R., & Rizzuti, B. (2016). Multiple binding modes of ibuprofen in human serum albumin identified by absolute binding free energy calculations. Physical Chemistry Chemical Physics, 18(47), 32358–32368. https://doi.org/10.1039/c6cp05680f
- Era, S., Kuwata, K., Imai, H., Nakamura, K., Hayashi, T., & Sogami, M. (1995). Age-related change in redox state of human serum albumin. Biochimica et Biophysica Acta (BBA) – Protein Structure and Molecular Enzymology, 1247(1), 12–16. https://doi.org/10.1016/0167-4838(94)00166-E
- Fan, J., Gilmartin, K., Octaviano, S., Villar, F., Remache, B., & Regan, J. (2022). Using human serum albumin binding affinities as a proactive strategy to affect the pharmacodynamics and pharmacokinetics of preclinical drug candidates. ACS Pharmacology & Translational Science, 5(9), 803–810. https://doi.org/10.1021/acsptsci.2c00115
- Fender, A. C., & Dobrev, D. (2019). Bound to bleed: How altered albumin binding may dictate warfarin treatment outcome. International Journal of Cardiology: Heart & Vasculature, 22, 214–215. https://doi.org/10.1016/j.ijcha.2019.02.007
- Fischer, N. H., Lopes van den Broek, S. I., Herth, M. M., & Diness, F. (2022). Radiolabeled albumin through SNAr of cysteines as a potential pretargeting theranostic agent. RSC Advances, 12, 35032–35036. https://doi.org/10.1039/D2RA06406E
- Fujiwara, S., & Amisaki, T. (2007). Identification of high-affinity fatty acid binding sites on human serum albumin by the MM-PBSA method. Biophysical Journal, 94(1), 95–103. https://doi.org/10.1529/biophysj.107.111377
- Ghuman, J., Zunszain, P. A., Petitpas, I., Bhattacharya, A. A., Otagiri, M., & Curry, S. (2005). Structural basis of the drug-binding specificity of human serum albumin. Journal of Molecular Biology, 353(1), 38–52. https://doi.org/10.1016/j.jmb.2005.07.075
- Gong, J., Yan, J., Forscher, C., & Hendifar, A. (2018). Aldoxorubicin: A tumor-targeted doxorubicin conjugate for relapsed or refractory soft tissue sarcomas. Drug Design, Development and Therapy, 12, 777–786. https://doi.org/10.2147/DDDT.S140638
- Green, M. R., Manikhas, G. M., Orlov, S., Afanasyev, B., Makhson, A. M., Bhar, P., & Hawkins, M. J. (2006). Abraxane, a novel cremophor-free, albumin-bound particle form of paclitaxel for the treatment of advanced non-small-cell lung cancer. Annals of Oncology, 17(8), 1263–1268. https://doi.org/10.1093/annonc/mdl104
- Harm, S., Schildböck, C., & Hartmann, J. (2018). Removal of stabilizers from human serum albumin by adsorbents and dialysis used in blood purification. PLoS ONE, 13(1), e0191741. https://doi.org/10.1371/journal.pone.0191741
- He, X. M., & Carter, D. C. (1992). Atomic structure and chemistry of human serum albumin. Nature, 358(6383), 209–215. https://doi.org/10.1038/358209a0
- Hoogenboezem, E. N., Patel, S. S., Lo, J. H., Cavnar, A. B., Babb, L. M., Francini, N., … Duvall, C. L. (2024). Structural optimization of siRNA conjugates for albumin binding achieves effective MCL1-targeted cancer therapy. Nature Communications, 15(1), 1581. https://doi.org/10.1038/s41467-024-45609-0
- Hoogenboezem, E. N., & Duvall, C. L. (2018). Harnessing albumin as a carrier for cancer therapies. Advanced Drug Delivery Reviews, 130, 73–89. https://doi.org/10.1016/j.addr.2018.07.011
- Johnson, J. A. (2012). Warfarin pharmacogenetics: A rising tide for its clinical value. Circulation, 125(16), 1964–1966. https://doi.org/10.1161/CIRCULATIONAHA.112.100628
- Kragh-Hansen, U. (1983). Relations between high-affinity binding sites for L-tryptophan, diazepam, salicylate, and phenol red on human serum albumin. Biochemical Journal, 209(1), 135–142. https://doi.org/10.1042/bj2090135
- Kragh-Hansen, U. (1991). Octanoate binding to the indole- and benzodiazepine-binding region of human serum albumin. Biochemical Journal, 273(3), 641–644. https://doi.org/10.1042/bj2730641
- Larsen, M. T., Kuhlmann, M., Hvam, M. L., & Howard, K. A. (2016). Albumin-based drug delivery: Harnessing nature to cure disease. Molecular and Cellular Therapies, 4, 3. https://doi.org/10.1186/s40591-016-0048-8
- Lau, J., Bloch, P., Schäffer, L., Pettersson, I., Spetzler, J., Kofoed, J., … Kruse, T. (2015). Discovery of the once-weekly glucagon-like peptide 1 (GLP-1) analogue semaglutide. Journal of Medicinal Chemistry, 58(18), 7370–7380. https://doi.org/10.1021/acs.jmedchem.5b00726
- Leblanc, Y., Berger, M., Seifert, A., Bihoreau, N., & Chevreux, G. (2019). Human serum albumin presents isoform variants with altered neonatal Fc receptor interactions. Protein Science, 28(11), 1982–1992. https://doi.org/10.1002/pro.3733
- Lee, P., & Wu, X. (2015). Modifications of human serum albumin and their binding effect. Current Pharmaceutical Design, 21(14), 1862–1865. https://doi.org/10.2174/1381612821666150302115025
- Liu, X., Mohanty, R. P., Maier, E. Y., Peng, X., Wulfe, S., Looney, A. P., Aung, K. L., & Ghosh, D. (2020). Controlled loading of albumin–drug conjugates ex vivo for enhanced drug delivery and antitumor efficacy. Journal of Controlled Release, 328, 1–12. https://doi.org/10.1016/j.jconrel.2020.08.015
- Loun, B., & Hage, D. S. (1994). Chiral separation mechanisms in protein-based HPLC columns. 1. Thermodynamic studies of (R)- and (S)-warfarin binding to immobilized human serum albumin. Analytical Chemistry, 66(21), 3814–3822. https://doi.org/10.1021/ac00093a043
- Madsen, B. K., Hilscher, M., Zetner, D., & Rosenberg, J. (2019). Adverse reactions of dimethyl sulfoxide in humans: A systematic review. F1000Research, 7, 1746. https://doi.org/10.12688/f1000research.16642.2
- Mehtala, J. G., Kulczar, C., Lavan, M., Knipp, G., & Wei, A. (2015). Cys34-PEGylated human serum albumin for drug binding and delivery. Bioconjugate Chemistry, 26(5), 941–949. https://doi.org/10.1021/acs.bioconjchem.5b00143
- Miele, E., Spinelli, G. P., Miele, E., Tomao, F., & Tomao, S. (2009). Albumin-bound formulation of paclitaxel (Abraxane® ABI-007) in the treatment of breast cancer. International Journal of Nanomedicine, 4, 99–105. https://doi.org/10.2147/ijn.s3061
- Mishra, V., & Heath, R. J. (2021). Structural and biochemical features of human serum albumin essential for eukaryotic cell culture. International Journal of Molecular Sciences, 22(16), 8411. https://doi.org/10.3390/ijms22168411
- Nagumo, K., Tanaka, M., Chuang, V. T. G., Setoyama, H., Watanabe, H., Yamada, N., Kubota, K., Tanaka, M., Matsushita, K., Yoshida, A., Jinnouchi, H., Anraku, M., Kadowaki, D., Ishima, Y., Sasaki, Y., Otagiri, M., & Maruyama, T. (2014). Cys34-cysteinylated human serum albumin is a sensitive plasma marker in oxidative stress-related chronic diseases. PLoS ONE, 9(1), e85216. https://doi.org/10.1371/journal.pone.0085216
- Oettl, K., & Marsche, G. (2010). Redox state of human serum albumin in terms of cysteine-34 in health and disease. In Methods in Enzymology (Vol. 474, pp. 181–195). Academic Press. https://doi.org/10.1016/S0076-6879(10)74011-8
- Olsen, H., Andersen, A., Nordbø, A., Kongsgaard, U. E., & Børmer, O. P. (2004). Pharmaceutical-grade albumin: Impaired drug-binding capacity in vitro. BMC Clinical Pharmacology, 4, 4. https://doi.org/10.1186/1472-6904-4-4
- O’Reilly, R. A. (1967). Studies on the coumarin anticoagulant drugs: Interaction of human plasma albumin and warfarin sodium. The Journal of Clinical Investigation, 46(5), 829–837. https://doi.org/10.1172/JCI105582
- Park, J. H., Lee, J. W., Kang, K., & Bae, S. H. (2021). Effect of stabilizers on drug binding capacity of human serum albumin. Pharmaceuticals, 14(12), 1295. https://doi.org/10.3390/ph14121295
- Petitpas, I., Bhattacharya, A. A., Twine, S., East, M., & Curry, S. (2001). Crystal structure analysis of warfarin binding to human serum albumin: Anatomy of drug site I. Journal of Biological Chemistry, 276(25), 22804–22809.
https://doi.org/10.1074/jbc.M100575200 - Rondeau, P., & Bourdon, E. (2011). The glycation of albumin: Structural and functional impacts. Biochimie, 93(4), 645–658. https://doi.org/10.1016/j.biochi.2010.12.003
- Ruggiero, A., Villa, C. H., Bander, E., Rey, D. A., Bergkvist, M., Batt, C. A., Manova-Todorova, K., Deen, W. M., Scheinberg, D. A., & McDevitt, M. R. (2010). Paradoxical glomerular filtration of carbon nanotubes. Proceedings of the National Academy of Sciences, 107(27), 12369–12374. https://doi.org/10.1073/pnas.0913667107
- Sand, K. M. K., Bern, M., Nilsen, J., Noordzij, H. T., Sandlie, I., & Andersen, J. T. (2015). Unraveling the interaction between FcRn and albumin: Opportunities for design of albumin-based therapeutics. Frontiers in Immunology, 5, 682.
https://doi.org/10.3389/fimmu.2014.00682 - Sarett, S. M., Werfel, T. A., Lee, L., Jackson, M. A., Kilchrist, K. V., Brantley-Sieders, D., & Duvall, C. L. (2017). Lipophilic iRNA targets albumin in situ and promotes bioavailability, tumor penetration, and carrier-free gene silencing. Proceedings of the National Academy of Sciences, 114(32), E6490–E6497. https://doi.org/10.1073/pnas.1621240114
- Scatchard, G. (1949). The attractions of proteins for small molecules and ions. Annals of the New York Academy of Sciences, 51(4), 660–672. https://doi.org/10.1111/j.1749-6632.1949.tb27297.x
- Spada, A., Emami, J., Tuszynski, J. A., & Lavasanifar, A. (2021). The uniqueness of albumin as a carrier in nanodrug delivery. Molecular Pharmaceutics, 18(5), 1866–1883. https://doi.org/10.1021/acs.molpharmaceut.1c00090
- Sudlow, G., Birkett, D. J., & Wade, D. N. (1975). The characterization of two specific drug-binding sites on human serum albumin. Molecular Pharmacology, 11(6), 824–832. https://pubmed.ncbi.nlm.nih.gov/1207674/
- Tramarin, A., Tedesco, D., Naldi, M., Baldassarre, M., Bertucci, C., & Bartolini, M. (2019). New insights into the altered binding capacity of pharmaceutical-grade human serum albumin: Site-specific binding studies by induced circular dichroism spectroscopy. Journal of Pharmaceutical and Biomedical Analysis, 162, 171–178. https://doi.org/10.1016/j.jpba.2018.09.022
- Vorum, H., Jørgensen, H. R., & Brodersen, R. (1993). Variation in the binding affinity of warfarin and phenprocoumon to human serum albumin in relation to surgery. European Journal of Clinical Pharmacology, 44(2), 157–162. https://doi.org/10.1007/BF00315474
- Ward, E. S., Gelinas, D., Dreesen, E., Van Santbergen, J., Andersen, J. T., Silvestri, N. J., Kiss, J. E., Sleep, D., Rader, D. J., Kastelein, J. J. P., Louagie, E., Vidarsson, G., & Spriet, I. (2022). Clinical significance of serum albumin and implications of FcRn inhibitor treatment in IgG-mediated autoimmune disorders. Frontiers in Immunology, 13, 892534. https://doi.org/10.3389/fimmu.2022.892534
- Weber, J. J., Kanost, M. R., & Gorman, M. J. (2020). Iron binding and release properties of transferrin-1 from Drosophila melanogaster and Manduca sexta: Implications for insect iron homeostasis. Insect Biochemistry and Molecular Biology, 125, 103438. https://doi.org/10.1016/j.ibmb.2020.103438
- Yamasaki, K., Maruyama, T., Yoshimoto, K., Tsutsumi, Y., Narazaki, R., Fukuhara, A., Kragh-Hansen, U., & Otagiri, M. (1999). Interactive binding to the two principal ligand-binding sites of human serum albumin: Effect of the neutral-to-base transition. Biochimica et Biophysica Acta (BBA) – Protein Structure and Molecular Enzymology, 1432(2), 313–323. https://doi.org/10.1016/S0167-4838(99)00098-9
- Yu, L., Hua, Z., Luo, X., Zhao, T., & Liu, Y. (2022). Systematic interaction of plasma albumin with the efficacy of chemotherapeutic drugs. Biochimica et Biophysica Acta (BBA) – Reviews on Cancer, 1877(1), 188655. https://doi.org/10.1016/j.bbcan.2021.188655
- Zhang, Y., Jia, Y., & Zhu, S. (2023). NIR-II cyanine@albumin fluorophore for deep tissue imaging and imaging-guided surgery. SmartMat, 4(1), e1245. https://doi.org/10.1002/smm2.1245
- Zhou, R., Zhong, R., Luo, Z., Wei, H., & Yu, C.-Y. (2025). Advances on albumin-based carriers for anticancer drug delivery. Advanced NanoBiomed Research, 5(9), 2500011. https://doi.org/10.1002/anbr.202500011