Transient Transfection Protocol – Enhancement of Transfection Efficiency Using Recombinant Transferrin with Serum-Free HEK293 Media
Published on 18 May 2020
Enhancement of Transient Transfection with Serum-Free and Blood-Free Transferrin
Abstract
Methods to enhance transient transfection of adherent cells have identified many different ligands that improve plasmid uptake into cells. Of these ligands, serum-derived transferrin is well-known to improve transfection efficiency of adherent cells. However, to date, a serum-free and blood-free transferrin has not been tested for improvements to transfection efficiency. We have thus created a protocol for the improvement of transfection efficiency and viral titer without the need for blood-derived transferrin. Excess transferrin does not negatively impact transfection, and the incorporation of a blood- and serum-free, recombinant human transferrin supports high transfection efficiency and viral titer in a chemically defined, blood-free medium.
Background: Improved Transient Transfection Protocol
Transient transfection relies on the spontaneous fusion of plasmid DNA with the cell membrane and the subsequent endocytosis and trafficking to the nucleus in order to achieve transient gene expression. Improvements made to polymers or lipids have resulted in numerous commercially available products for highly efficient transfection. However, these products are still subject to fusion with the cells and the ability of cells to endocytose foreign DNA.
Methods to improve a cell’s ability to take in plasmid DNA have identified that transmembrane receptors can serve as ports of entry for the internalization of extracellular DNA. While there are thousands of receptors that can be present on a cell, the transferrin receptor 1 (TfR1) is ubiquitously expressed, as every cell requires extracellular iron for health and growth. Transferrin has been identified as a therapeutic agent for the delivery of nanoparticles and small molecules due to the ability of transferrin to be covalently linked and also as a natural carrier of molecules for entering a cell through its receptors.1
Given the success of transferrin as a molecular carrier, the use of transferrin has still relied on serum-derived transferrin from either bovine serum or human serum. Both of which are subject to numerous disadvantageous factors such as adventitious agents present in serum, constraints on the supply-chain, or increasing cost due to demand. We have therefore developed a protocol for the use of a recombinant human transferrin that is not derived from serum or animal blood.
While excess transferrin in transfection has already been identified to increase transfection efficiency,2 we show that blood-free and serum-free transferrin added in excess during the transfection of adherent cells improves the transfection efficiency compared to low levels of transferrin and serum-derived transferrin. We also show that the inclusion of transferrin in transient transfection for viral production enhanced functional viral titers for lentivirus. Our data suggest that serum-free and blood-free transferrin can be used as a carrier for nucleic acids and potentially other molecules for trafficking molecules into cells.
Methods
Cell Lines and Culture
HEK-293T (293T, ATCC CRL-3216) and HT-1080 (ATCC CCL-121) were maintained in either DMEM + 10% FBS for serum conditions, or complete OptiPEAK HEK293t® (InVitria, Junction City, Kansas), for serum-free conditions. Serum cultured cells were grown in standard conditions (5% CO2, 37 °C) while serum-free cells were grown in 10% CO2 and 37 °C. Cells were adapted to serum-free conditions by direct adaptation from FBS passaged cells. Both cell types were maintained by passaging with TrypLE (ThermoFisher #A1217701) every 3-4 days and seeded at initial densities of 10,000 cells/cm2. 293T cells were adapted to serum-free conditions over three passages.
Transfection and Reagents
293T cells for transfection were plated 24 hours prior to transfection at a density of 50,000 cells per cm2 for GFP efficiency assays or for lentivirus production cells were plated at a density of 75,000 cells per cm2. For transfection efficiency with a GFP reporter, the plasmid Monster Green® Fluorescent Protein phMGFP Vector (Promega, Madison, WI) was used at a concentration of 0.2 µg/cm2. Plasmids for 2nd generation lentiviral vector production included: psPAX2 (Addgene plasmid #12260), pMD2.G (Addgene plasmid #12259), and pSIH1-H1-siLuc-copGFP transfer plasmid (System Biosciences, Palo Alto, CA).
Lentivirus plasmids were delivered at a concentration of 0.16 µg/cm2. In all cases, PEIpro (Polyplus, Alsace, France) was used at a ratio of 1:1 with plasmid DNA for formation of DNA complexes. For serum-free conditions, complexes were formed in complete OptiPEAK HEK293t and for serum conditions complexes were formed with OptiMEM (ThermoFisher, #31985070).
Transferrin supplementation was carried out by adding reconstituted Optiferrin® (InVitria, Junction City, Kansas) to complete OptiPEAK HEK293t at a final concentration of 1 mg/mL. To form complexes, plasmid DNA and PEIpro were first diluted separately 40-fold. Diluted DNA was then added to diluted PEIpro, the contents were mixed by gentle pipetting or swirling, and complexes were formed under sterile conditions for 15-20 minutes at room temperature. The reaction volume was then adjusted to a final volume of 0.08 mL/cm2, and the entire contents were added to the cell monolayer following aspiration of the conditioned media.
Cells were incubated in normal growth conditions for 2.5 hours ± 30 minutes, after which transfection complexes were aspirated off of cells, and normal growth media were added back to cells (without additional transferrin for transferrin enhancement). Cells were grown for 48 hours and then harvested for analysis by flow cytometry (Accuri C6 plus, BD, Palo Alto, California) or viral titering.
Functional Viral Titering
HT-1080 cells were used for functional transduction assays and were plated at a density of 30,000 cells per well in 12 well plates two hours before the assay. Lentivirus viral particles were harvested by collecting the total supernatant 48 hours post-transfection. The supernatant was briefly centrifuged for five minutes at room temperature to clear cell debris, and then titered by as previously described in the presence of Polybrene.3
Results and Conclusions
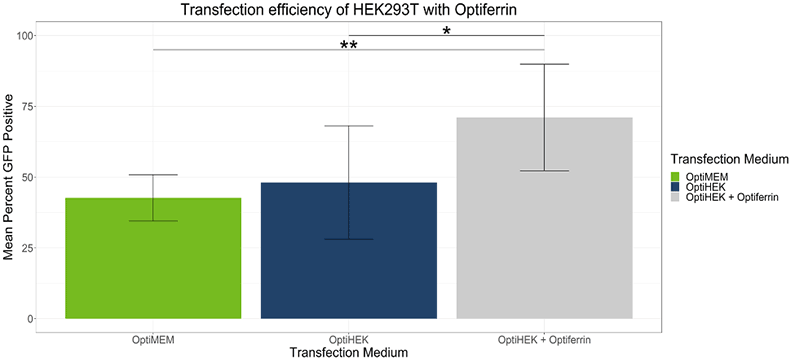
In standard conditions, OptiPEAK HEK293t achieves equivalent transfection efficiency compared to transfections performed under serum conditions complexed with OptiMEM. The average percentage of GFP positive cells complexed with OptiMEM and grown in DMEM + 10% FBS was 42.6% ± 8.1% and the average with OptiPEAK HEK293t was 48.1% ± 20.1% (Figure 1, difference in means is not statistically significant). Including excess transferrin in the transfection medium of OptiPEAK HEK293t raised the average percent GFP positive cells significantly to 71.1% ± 18.8% compared to DMEM + 10% FBS and OptiPEAK HEK293t without excess transferrin (Figure 1, p < 0.05 by Student’s t-test).
These results indicate that excess transferrin does not negatively affect transfection and can enhance transfection efficiency of HEK-293T cells in serum-free, chemically defined conditions.
To determine if transferrin enhancement of transfection efficiency correlated with higher viral titers, we performed transient transfection with lentivirus plasmids for viral production with 293T cells. Excess transferrin in the transfection step for viral production led to an increase in functional viral titer for lentivirus compared to Complete OptiPEAK HEK293t without excess transferrin (Figure 2). Lentivirus production showed similar trends to transfection efficiency assays with a GFP reporter. Second generation lentivirus vectors were produced using a three plasmid system, and total lentivirus vectors were harvested 48 hours post transfection. Lentivirus plasmids complexed with OptiMEM or OptiPEAK HEK293t showed near equivalent functional titers, with the average titer from plasmids complexed with OptiMEM and then cells grown with DMEM + 10% FBS was 1.79 ± 1 × 107 IFU/mL (Infectious Units/mL) (Figure 2).
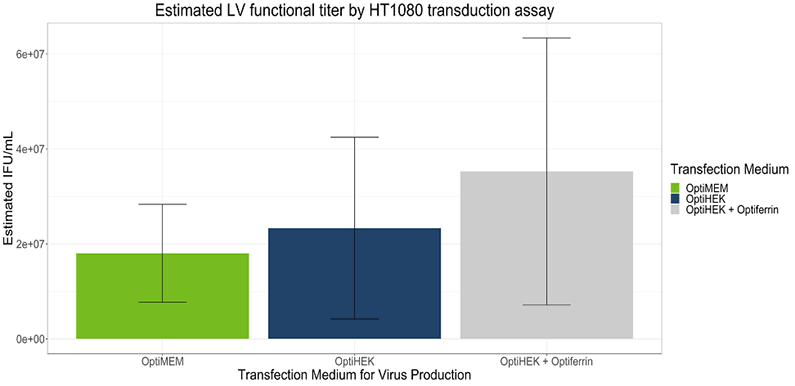
Plasmids complexed with OptiPEAK HEK293t and grown in chemically defined conditions with OptiPEAK HEK293t yielded average titer of 2.3 ± 1.9 × 107 IFU/mL (Figure 2, difference in means is not statistically significant). Plasmids complexed with excess transferrin in OptiPEAK HEK293t showed a log-fold increase over OptiPEAK HEK293t in average functional titer, with an average of 3.53 ± 2.8 × 107 IFU/mL (Figure 2).
Together, these results demonstrate that excess transferrin does not inhibit DNA complexation and in fact improves functional lentivirus titer compared to chemically defined conditions without excess transferrin and improves functional titer compared to serum-derived transferrin in OptiMEM. The improvements shown here with excess transferrin suggest that 293T cells may be more rapidly endocytosing plasmid DNA from the complexation medium. Another hypothesis for the improvements shown here is that the plasmid DNA is better protected within the cell as it is trafficked to the nucleus.
Regardless of the mechanism, a serum-free, blood-free, recombinant human transferrin is able to achieve high transfection efficiency, and high viral titer, suggesting that blood-free transferrin has high functionality in a chemically-defined medium.
The following content is gated. Please, subscribe to open access to it.
Footnotes
- Daniels, T. R., et al. (2012). “The transferrin receptor and the targeted delivery of therapeutic agents against cancer.” Biochim Biophys Acta 1820(3): 291-317.
- Tros de Ilarduya, C. and N. Duzgunes (2013). “Delivery of therapeutic nucleic acids via transferrin and transferrin receptors: lipoplexes and other carriers.” Expert Opin Drug Deliv 10(11): 1583-1591.
- Sena-Esteves, M. and G. Gao (2018). “Titration of Lentivirus Vectors.” Cold Spring Harb Protoc 2018(4).