Functional Characterization of Lacromin® Recombinant Human Holo-Lactoferrin for Cell Culture and Advanced Therapy Applications
Published on 25 June 2026
Application Note
Executive Summary
Here we compare Lacromin, InVitria’s recombinant human iron-saturated lactoferrin, to human milk-derived LF (hLF) and bovine milk-derived LF (bLF) in three functional assays: iron binding, cellular uptake by Caco-2 intestinal epithelial cells, and proliferation of human intestinal epithelial crypt-like cells (HIEC). Lacromin matched the functional performance of hLF in all three assays and showed greater uptake by human intestinal cells than bLF. These results, combined with the breadth of lactoferrin’s reported biological activities, such as wound healing, neuroprotection, and immunomodulation, support Lacromin as a functionally active, animal-origin-free lactoferrin for cell culture, translational research, and advanced therapy development.
Key Highlights
- Functionally equivalent to human milk-derived lactoferrin
- Greater human cellular uptake than bovine-derived lactoferrin
- Iron-saturated (holo) form for consistent biological activity
- Facilitates AOF, cGMP cell and gene therapy workflows
Introduction
Lactoferrin is one of the most abundant proteins in mammalian milk, with human milk containing some of the highest LF concentrations across species (Rai et al., 2014). LF’s primary function is to bind iron to control iron delivery while moderating oxidative stress, a property that underlies many of its reported biological activities including immune regulation, antimicrobial defense, neuroprotection, wound healing, and intestinal epithelial function. LF is critical for proper neonatal development of the brain, the gastrointestinal tract, and the innate immune system. Moreover, the gut-brain axis links intestinal LF activity between the gut, immune, and neurodevelopment through shared iron handling, immune signaling, and microbiota-mediated effects (Donovan, 2016).
Infants who are deficient in nutritional needs such as lactoferrin are associated with increased risk of infection, necrotizing enterocolitis, and neonatal sepsis (Schirmbeck et al., 2022). LF supports multiple aspects of healthy infant development, including intestinal immune homeostasis, gut barrier maturation, and cognitive development (Donovan, 2016). In fact, a randomized controlled trial showed that infants receiving formula supplemented with bovine milk fat globule membrane and LF for 12 months had higher neurodevelopmental scores at 5.5 years of age, including a 5-point IQ increase relative to control formula (Colombo et al., 2023). Furthermore, LF has also been studied for clinical applications including neonatal sepsis prevention, gastrointestinal therapeutics, immune support, wound care, and adjunctive cancer therapy (Manzoni, 2016; Wolf et al., 2007). Beyond its central role in infant development, LF is a multifunctional regulator of adult host physiology, with biological activities spanning iron sequestration, antimicrobial defense, immune signaling, neuroprotection, wound healing, and modulation of oxidative stress (Rizzi et al., 2025; Shini et al., 2022). Many of these same properties are also relevant to cell culture, regenerative medicine, and advanced therapy development, where iron homeostasis, cellular stress responses, and immune regulation influence cell function and manufacturing outcomes.
LF’s iron-binding activity underlies many of its functions, including bacteriostatic effects in the gut and at mucosal surfaces. The iron saturation of LF is a critical determinant of its biological activity, influencing receptor interactions and signaling. LF exists in two forms: holo-LF (iron-saturated) and apo-LF (iron-free). In intestinal epithelial cells, these forms activate distinct signaling pathways: holo-LF primarily signals through PI3K/Akt, while apo-LF more strongly activates ERK1/2 (Liao et al., 2012). This pathway-level difference matters in practice because it influences which downstream cellular responses are engaged. PI3K/Akt drives cell survival, growth, and metabolic signaling, while ERK1/2 drives proliferation and mitogenic cues, meaning the iron-saturation state of an LF preparation is not a passive property but a factor in biological outcome. Lacromin is supplied in the holo form, ensuring consistent iron-saturated activity across applications. This is particularly important because the biological effects of lactoferrin can vary depending on its iron saturation state.
Most commercial LF supplementation today uses bovine-derived material, while clinical and food-grade use of recombinant human LF has been more limited. Recombinant human LF retains functional activity, including the induction of antioxidant enzymes (Kruzel et al., 2013), and offers practical advantages for regulated applications: consistent supply, defined composition, and no animal-origin material (Vishwanath-Deutsch et al., 2024). Recent work has also shown that fermentation-derived recombinant bovine LF is functionally comparable to native bovine LF in intestinal cell assays (Skoog et al., 2025), supporting the broader proposition that recombinant LF can serve as a functional substitute for milk-derived material.
This application note surveys the major application areas where Lacromin’s defined composition and human-sequence holo form are most relevant, with attention to whether the available evidence is from human or animal LF and from holo or apo forms. We then evaluated Lacromin (InVitria’s recombinant human holo-LF) against human and bovine milk–derived LF in three assays representing core LF activities: iron binding, uptake by intestinal epithelial cells, and induction of intestinal epithelial cell proliferation. These assays were chosen to test whether a recombinant human holo-LF retains the functional behavior expected of native milk-derived human LF in a defined epithelial system, providing the experimental foundation for the application areas described.
Potential Applications of Lactoferrin in Cell Culture and Advanced Therapy Research
The sections below cover each application area in turn, summarizing the underlying biology and key studies.
Lactoferrin in Regulatory T Cell (Treg) Therapy Research
Treg therapies represent a rapidly expanding class of advanced therapeutics for autoimmune diseases, transplant rejection, and inflammatory disorders. Manufacturing of functional Tregs requires ex vivo expansion conditions that support Foxp3 expression, suppressive capacity, and phenotypic stability.
Recombinant human lactoferrin has been shown to promote Foxp3+ Treg differentiation and reduce inflammation in murine models of inflammatory bowel disease (MacManus et al., 2017). Lactoferrin combined with TGF-β1 and retinoic acid robustly generates inducible Tregs (iTregs) with potent suppressive function (Jang et al., 2023). Lactoferrin also influences the upstream antigen-presenting environment: LF-treated dendritic cells adopt a tolerogenic phenotype with enhanced capacity to drive Foxp3+ iTreg differentiation (Park et al., 2020).
These findings suggest that lactoferrin may serve as a functional supplement in Treg expansion and differentiation protocols, providing Foxp3-promoting activity alongside iron delivery and immunomodulatory support.
Lactoferrin in Neuroprotection and Parkinson’s Disease Research
Neurodegenerative diseases, particularly Parkinson’s disease (PD), are characterized by progressive loss of dopaminergic neurons driven in part by iron-mediated oxidative stress and neuroinflammation (Li and Guo, 2021). Lactoferrin combines high-affinity iron binding with the ability to cross the blood-brain barrier via receptor-mediated transcytosis (Li and Guo, 2021). In MPTP-treated mouse models of PD, lactoferrin administration ameliorated dopaminergic neurodegeneration and motor deficits by suppressing iron accumulation, upregulating brain-derived neurotrophic factor (BDNF), and activating the ERK/CREB signaling pathway (Xu et al., 2018), all of which are important in neurodevelopment and cognition. MPTP is a neurotoxin that experimentally induces PD-like motor dysfunction. Pretreatment with human lactoferrin partially preserved tyrosine hydroxylase-positive neurons and improved motor function after acute MPTP exposure (Kopaeva et al., 2021). The endogenous role of LF in neuroprotection has also been demonstrated: astrocytic lactoferrin deficiency worsened MPTP-induced dopaminergic neuron loss (Xu et al., 2024). Lactoferrin has also been proposed as a candidate therapeutic for synucleinopathies (Barros and Vieira, 2025).
The antioxidant properties of LF, including induction of superoxide dismutase and glutathione peroxidase, further support neuronal survival under oxidative stress (Kruzel et al., 2013; Rascón-Cruz et al., 2025). For researchers developing neuroprotective compound screens, iPSC-derived dopaminergic neuron models, or iron biology studies in the central nervous system, Lacromin provides a controlled, human-sequence, iron-saturated tool for these applications.
Lactoferrin in Wound Healing and Skin Biology
Wound healing is a coordinated, multi-phase process involving hemostasis, inflammation, granulation tissue formation, re-epithelialization, and tissue remodeling. Lactoferrin participates in multiple phases (Takayama and Aoki, 2012). During inflammation, LF modulates neutrophil and macrophage activity and suppresses excess pro-inflammatory cytokine release (Conesa et al., 2023). Its antimicrobial activity simultaneously protects wound sites from infection, including antibiotic-resistant pathogens such as methicillin-resistant Staphylococcus aureus (Kosznik-Kwaśnicka et al., 2026).
Rice-derived recombinant human LF stimulates human dermal fibroblast proliferation, migration, and survival under serum-free conditions, with effects mediated through MEK/ERK, Src, and Rho/ROCK signaling (Tang et al., 2010a). The same recombinant LF stimulates human keratinocyte proliferation, migration, and wound re-epithelialization in scratch-wound assays (Tang et al., 2010b). During remodeling, LF supports extracellular matrix deposition, including collagen and hyaluronan synthesis (Takayama and Aoki, 2012).
In dermatological applications, LF enhances transepithelial electrical resistance in reconstructed human epidermal models, inhibits pro-inflammatory mediators, and upregulates barrier proteins including aquaporin-3, filaggrin, and involucrin (Xie et al., 2024). Bovine LF has also been shown to promote dermal papilla cell proliferation and hair growth via ERK/Akt and Wnt signaling (Huang et al., 2019).
Lactoferrin as an Antioxidant and Anti-Apoptotic Supplement for Cell Expansion
Ex vivo cell expansion, particularly of mesenchymal stem cells (MSCs), T cells, and iPSC-derived lineages, is often challenged by oxidative stress, replicative senescence, and apoptosis under serum-free or low-density conditions. Defined, animal-origin-free supplements that mitigate these stresses have direct relevance to advanced therapy manufacturing.
Lactoferrin has been shown to protect human MSCs from oxidative stress-induced senescence and apoptosis (Park et al., 2017). Consistent anti-apoptotic effects have been demonstrated in rat bone marrow mesenchymal stem cells, where LF reduces apoptosis while promoting proliferation and directing differentiation toward osteogenic over adipogenic lineages (An et al., 2019). In primary human skin fibroblasts, rice-derived holo-rhLF protects against apoptosis under serum-free conditions as measured by TUNEL and calcein-AM/PI staining, and sustains cell survival alongside its proliferative effects (Tang et al., 2010a). LF also modulates the differentiation of pluripotent mesenchymal cells, promoting osteogenic and chondrogenic lineage commitment via LRP1 (Yagi et al., 2009).
Mechanistically, lactoferrin’s antioxidant activity operates through both direct iron sequestration and induction of endogenous antioxidant defense, including upregulation of superoxide dismutase (SOD), glutathione peroxidase (GPX), and Nrf2-dependent gene expression (Kruzel et al., 2013; Rascón-Cruz et al., 2025). Anti-cellular senescence effects have also been documented in aging-focused reviews (Li et al., 2021).
Rice-derived recombinant human LF has established utility as a cell culture supplement, with demonstrated effects on the growth of intestinal, hybridoma, osteoblast, and human embryonic kidney cell lines (Huang et al., 2008). LF can also localize to the nucleus of intestinal epithelial cells following receptor-mediated uptake (Suzuki et al., 2008), where it binds DNA with sequence specificity (He and Furmanski, 1995) and has been associated with modulation of immune and tissue-repair gene transcription (Lönnerdal et al., 2020; Kruzel et al., 2013).
Direct studies of recombinant human holo-LF in human MSC expansion remain limited. Nevertheless, across diverse cell types, Lacromin is suitable for serum-free media optimization, defined cell expansion workflows, and cGMP-aligned cell and gene therapy manufacturing where raw-material consistency and traceability are required (Vishwanath-Deutsch et al., 2024).
Lactoferrin in Cancer Research and Tumor Biology
Lactoferrin has emerged as a multi-mechanism modulator of tumor biology, with reported effects on apoptosis, ferroptosis, angiogenesis, and the hypoxic tumor microenvironment. Recent reviews summarize converging mechanisms across cancer types, including iron metabolism dysregulation, immune modulation, and modulation of apoptosis and ferroptosis (Gallo and Antonini, 2024; Bolesławska et al., 2025).
Iron-saturation state can influence LF’s role and activity in cancer models. In a study of triple-negative breast cancer (TNBC), holo-lactoferrin specifically released labile iron, promoted reactive oxygen species generation via Fenton chemistry, and induced ferroptosis in MDA-MB-231 cells. The same iron-driven mechanism allowed holo-LF to catalyze H₂O₂ decomposition, ameliorate the hypoxic tumor microenvironment, and downregulate HIF-1α. In a nude mouse xenograft model, holo-LF combined with 4 Gy radiation enhanced anti-tumor efficacy and improved survival relative to radiation alone. By contrast, apo-LF was not cytotoxic and inhibited ferroptosis (Zhang et al., 2021).
These effects are cell-type dependent: non-TNBC MCF-7 cells were resistant to holo-LF-induced ferroptosis owing to their higher redox and iron-handling capacity (Zhang et al., 2021). Researchers should therefore validate LF activity in their specific cancer model rather than assuming a uniform anticancer effect across tumor types.
LF is also under active investigation as a tumor-targeting drug delivery vehicle, leveraging LF receptor expression on cancer cells and its ability to cross the blood-brain barrier for CNS tumor applications (Elzoghby et al., 2020; Pan et al., 2021).
For investigators studying tumor iron biology, ferroptosis, or radiosensitization, or developing LF-based drug delivery systems, Lacromin provides a defined, iron-saturated, human-sequence research tool aligned with the holo-LF mechanism described in the published literature.
Results and Discussion
The application areas surveyed above all depend on a small set of core lactoferrin functions: iron binding, receptor-mediated uptake into target cells, and induction of cellular responses such as proliferation. To evaluate whether Lacromin retains these core functions in a defined, recombinant, human-sequence form, we tested it against bovine and human milk-derived LF in these assays representing each: iron saturation and reversible iron binding, uptake by Caco-2 intestinal epithelial cells, and induction of human intestinal epithelial crypt-like cells (HIEC) proliferation.
Lacromin Demonstrates Reversible Iron Binding and Release
Human LF is an iron-binding glycoprotein containing two lobes (N and C), each capable of binding one ferric iron ion. Lactoferrin binds iron with high affinity and can reversibly release it under acidic conditions. Structural studies show that iron binding and release involve conformational transitions between open (apo) and closed (holo) states (Baker and Baker, 2004). In its native form, hLF is partially saturated with iron (5%–30%), but it can also exist in apo-LF form characterized by low iron (<5%) or holo-LF form characterized by high iron (>70%) (Rizzi et al., 2025). The degree of iron binding is typically reported as a degree of saturation, taking into account that full iron saturation (100%) means binding of two ferric iron molecules per one LF molecule. The iron saturation state of lactoferrin is a critical determinant of its biological activity, influencing receptor interactions, signaling pathway activation, and downstream cellular responses (Liao et al., 2012; Zhang et al., 2021).
Table 1/Figure 1 shows that at baseline, Lacromin lots have high iron saturation (>70%), consistent with holo-LF, whereas human milk LF has low saturation, consistent with an apo-like state. Under acidic conditions (pH ~2), all samples release iron efficiently, reducing saturation to near zero. Upon return to neutral pH with available iron, all samples regain high levels of iron saturation, demonstrating preserved binding functionality. This reversible behavior is consistent with published findings showing that lactoferrin releases iron at low pH and can be fully re-saturated if structural and functional integrity is maintained.
Iron Binding and Release Characteristics of Lactoferrin — % Iron Saturation
| Samples | Baseline | Iron Released | Iron Saturated |
|---|---|---|---|
| Lacromin Lot 1 | 77.5 | 0.9 | 93.8 |
| Lacromin Lot 2 | 78.1 | 1.3 | 90.2 |
| Lacromin Lot 3 | 91.6 | 6.1 | 90.7 |
| Human milk LF | 6.2 | -0.1 | 88.2 |
| Bovine LF | 74.4 | -0.3 | 58.6 |
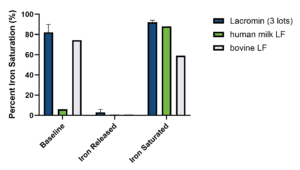
Table 1/Figure 1. Iron saturation and pH-dependent iron release/rebinding of lactoferrin samples. Lacromin lots showed high baseline iron saturation, released iron under acidic conditions, and recovered high saturation after iron reloading.
Overall, Lacromin demonstrates a pH-dependent iron-binding profile qualitatively comparable to human milk LF and bovine LF, indicating functional similarity in iron-binding and release.
Lacromin Matches Native Human LF in Caco-2 Uptake
Caco-2 cells are a widely used cancer cell model of intestinal epithelial barrier function and are often represented in applications of oral drug absorption alongside HIEC (Hidalgo et al., 1989; Takenaka et al., 2014). To demonstrate cellular internalization of LF, Caco-2 cells were treated with hLF, bLF, or Lacromin at 50 µg/mL at 37°C for one hour, and uptake was quantified by intracellular flow cytometry using polyclonal anti-lactoferrin FITC conjugated antibodies. Lacromin and hLF showed comparable uptake, with no statistically significant difference between them (Figure 2). To confirm that LF was internalized rather than simply surface-adsorbed, an unfixed control sample was also tested, and no staining was observed (data not shown). Bovine LF was taken up by fewer cells. These observations are consistent with prior reports comparing bovine and human LF binding to human intestinal cells (Lönnerdal et al., 2011; Jiang et al., 2014) and are likely due to structural differences between bovine and human lactoferrin. Bovine LF differs from human LF in approximately 31% of its amino acid sequence (Jiang et al., 2014), glycosylation architecture (Zlatina and Galuska, 2020), and receptor binding properties (Lönnerdal et al., 2011). These differences can affect cellular uptake efficiency and downstream biological responses in human systems (Jiang and Lönnerdal, 2014). Commercial bLF preparations also vary in iron saturation and lot-to-lot consistency (Lönnerdal et al., 2020), in contrast to the defined iron-saturation state of Lacromin.
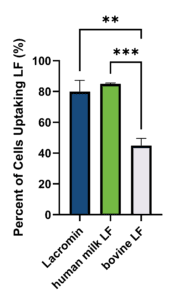
Figure 2. Uptake of LF by Caco-2 cells. Caco-2 cells were treated with 50 µg/mL of hLF, Lacromin, or bLF for 1 hour at 37°C. Internalized LF was detected by FITC-conjugated anti-lactoferrin antibody and quantified by intracellular flow cytometry as the percentage of FITC-positive cells. Data are mean ± SD; n = 3 biological replicates. ** p < 0.01, ***p < 0.001 by ordinary one-way ANOVA with Tukey post hoc test.
Lacromin Matches Native Human LF in HIEC Proliferation
LF promotes proliferation of intestinal epithelial cells, contributing to mucosal growth and barrier maintenance (Buccigrossi et al., 2007; Lönnerdal et al., 2011). To test whether Lacromin retains activity under conditions supporting both ERK1/2 and PI3K/Akt pathways, HIECs were treated with hLF, bLF, or Lacromin at 200 µg/mL in serum-free, EGF-supplemented medium for 48 hours, with proliferation measured by EdU incorporation. EGF was retained throughout the assay to support baseline ERK1/2 signaling, since holo-LF primarily activates PI3K/Akt rather than ERK1/2 in intestinal epithelial cells (Liao et al., 2012). Lacromin and hLF produced equivalent levels of HIEC proliferation, with no statistically significant difference between them; bLF produced similar proliferation to the human LFs (Figure 3). These results are consistent with prior comparability work showing that recombinant LFs retain the proliferative activity of native milk-derived LFs in intestinal cell assays (Skoog et al., 2025; Jiang et al., 2014).
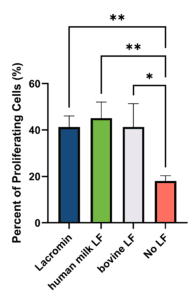
Figure 3. Effect of LF on HIEC proliferation. HIECs were seeded at 9,000 cells/well in 96-well plates, cultured for 24 hours, and serum-starved for 24 hours. Cells were then treated with hLF, bLF, or Lacromin at 200 µg/mL in serum-free medium for 48 hours, with EdU added during the final 24 hours. Proliferation was quantified by Click-iT EdU assay and reported as % EdU-positive cells. Data are mean ± SD; n = 3 biological replicates. *p < 0.05, **p < 0.01 by ordinary one-way ANOVA with Tukey post hoc test.
The comparable HIEC proliferation across all three LFs, despite the lower Caco-2 uptake of bLF, likely reflects the different concentrations and time windows of the two assays and saturation of the proliferative signaling response.
Conclusion
Lacromin performs comparably to human milk LF in iron binding, uptake by intestinal epithelial cells, and HIEC proliferation. Bovine LF showed lower uptake in Caco-2 cells, consistent with species differences in LF receptor binding, while producing similar HIEC proliferation under EGF-supplemented conditions. These results support Lacromin as a functional, animal-origin-free alternative to native human LF where a chemically defined, human-sequence lactoferrin is required.
Beyond these specific assays, the breadth of LF’s reported biological activities maps onto several areas of active therapeutic development. Its capacity to promote regulatory T cell differentiation, support mesenchymal stem cell survival and expansion, protect cells from oxidative and apoptotic stress, and serve as a tumor-targeting delivery scaffold positions Lacromin as a candidate raw material across cell therapy, regenerative medicine, and translational research. For cell and gene therapy manufacturing in particular, where supply consistency, defined composition, and the absence of animal-origin material are increasingly required, Lacromin offers a single human-sequence lactoferrin suited to evaluation across these applications.
Featured Solution
Lacromin – Recombinant Human Holo-Lactoferrin – Animal-Origin-Free, GMP-Produced
Lacromin is a chemically defined, recombinant human holo-lactoferrin (iron-saturated) designed to provide defined lactoferrin supplementation for cell culture and advanced therapy workflows. In this application note, Lacromin matched the functional performance of human milk-derived lactoferrin across three core assays: reversible iron binding and release, uptake by Caco-2 intestinal epithelial cells, and induction of HIEC proliferation. It also showed greater human cellular uptake than bovine-derived lactoferrin. Supplied in the holo form, Lacromin delivers consistent iron-saturated activity, a critical determinant of lactoferrin’s receptor interactions, signaling, and downstream cellular responses.
Unlike bovine milk-derived lactoferrin, Lacromin is animal-origin-free and human-sequence, avoiding the species-related sequence and glycosylation differences, lot-to-lot iron-saturation variability, and supply and regulatory concerns associated with milk-derived material. As a chemically defined recombinant protein, Lacromin integrates into serum-free media optimization, defined cell expansion workflows, and cGMP-aligned cell and gene therapy manufacturing where raw-material consistency and traceability are required.
Lacromin is well suited for cell therapy, gene therapy, regenerative medicine, and translational research workflows, including Treg expansion, mesenchymal stem cell survival and expansion, neuroprotection screens, and tumor-biology research, where a defined, human-sequence, iron-saturated lactoferrin is critical.
Frequently Asked Questions (FAQs)
What problem does this application note address? This application note addresses the need for a defined, animal-origin-free lactoferrin for cell culture and advanced therapy development. Most commercial lactoferrin (LF) is bovine milk-derived, which differs from human LF in sequence, glycosylation, and receptor binding, and varies in iron saturation and lot-to-lot consistency. The note evaluates whether Lacromin, InVitria’s recombinant human holo-lactoferrin, retains the functional behavior of native human milk-derived LF in a defined system, providing a human-sequence, chemically defined alternative for regulated workflows.
How was Lacromin evaluated in this study? Lacromin was compared against human milk-derived LF (hLF) and bovine milk-derived LF (bLF) in three assays representing core lactoferrin functions: iron saturation and reversible iron binding, uptake by Caco-2 intestinal epithelial cells, and induction of human intestinal epithelial crypt-like cell (HIEC) proliferation. These assays were chosen to test whether a recombinant human holo-LF behaves like native milk-derived human LF in a defined epithelial system.
What did the iron binding assay show? Lacromin demonstrated reversible, pH-dependent iron binding consistent with the holo form. At baseline, Lacromin lots showed high iron saturation (greater than 70%), compared with human milk LF, which showed low saturation consistent with an apo-like state. Under acidic conditions (pH approximately 2), all samples released iron to near zero, and upon return to neutral pH with available iron, all samples regained high saturation, demonstrating preserved binding functionality. Overall, Lacromin’s iron-binding profile was qualitatively comparable to both human and bovine milk LF.
What did the Caco-2 uptake assay show? Lacromin and hLF showed comparable cellular uptake, with no statistically significant difference between them. Bovine LF was taken up by fewer cells. This is consistent with prior reports comparing bovine and human LF binding to human intestinal cells and is likely due to structural differences between the two, including an approximately 31% amino acid sequence difference, distinct glycosylation, and different receptor binding properties.
What did the HIEC proliferation assay show? Lacromin and hLF produced equivalent levels of HIEC proliferation, with no statistically significant difference between them, and bLF produced similar proliferation to the human LFs. The assay was run in serum-free, EGF-supplemented medium to support baseline ERK1/2 signaling, since holo-LF primarily activates PI3K/Akt rather than ERK1/2 in intestinal epithelial cells. These results are consistent with prior work showing recombinant LFs retain the proliferative activity of native milk-derived LFs.
Why does the iron-saturation (holo) form matter? The iron saturation state of lactoferrin is a critical determinant of its biological activity, influencing receptor interactions and signaling. In intestinal epithelial cells, holo-LF (iron-saturated) primarily signals through PI3K/Akt, while apo-LF (iron-free) more strongly activates ERK1/2. Because these pathways drive different downstream responses, iron saturation is a functional property rather than a passive one. Lacromin is supplied in the holo form, providing consistent iron-saturated activity across applications.
Why use Lacromin instead of bovine or milk-derived lactoferrin? Lacromin is a recombinant, animal-origin-free, human-sequence lactoferrin. It avoids the species-related sequence and glycosylation differences of bovine LF, the lot-to-lot iron-saturation variability of commercial bovine preparations, and the supply and regulatory concerns associated with milk-derived material. Recombinant human LF retains functional activity and offers consistent supply, defined composition, and no animal-origin material, making Lacromin suitable for serum-free media optimization, defined cell expansion, and cGMP-aligned cell and gene therapy manufacturing where raw-material consistency and traceability are required.
What applications is Lacromin relevant to? Beyond the assays reported here, the breadth of lactoferrin’s documented biological activities maps onto several areas of active development, including regulatory T cell (Treg) expansion, mesenchymal stem cell survival and expansion, neuroprotection and Parkinson’s disease research, wound healing and skin biology, and cancer and tumor biology research. Lacromin provides a defined, human-sequence, iron-saturated tool for evaluation across cell therapy, gene therapy, regenerative medicine, and translational research, with the note flagging throughout whether supporting evidence derives from human or animal LF and from holo or apo forms.
The following content is gated. Please, subscribe to open access to it.
Footnotes
References
- Actor, J.K., Hwang, S.-A., & Kruzel, M.L. (2009). Lactoferrin as a natural immune modulator. Current Pharmaceutical Design, 15(17), 1956–1973. https://doi.org/10.2174/138161209788453202
- An, J., Xu, Y., Kong, Z., Xie, Y., Tabys, D., Ma, M., Cao, X., Ren, H., & Liu, N. (2019). Effect of lactoferrin and its digests on differentiation activities of bone mesenchymal stem cells. Journal of Functional Foods, 57, 202–210. https://doi.org/10.1016/j.jff.2019.04.020
Baker, H. M., & Baker, E. N. (2004). Lactoferrin and Iron: Structural and dynamic aspects of binding and release. Biometals, 17(3), 209–216. https://doi.org/10.1023/B:BIOM.0000027694.40260.70 - Barros, C. A., & Vieira, T. C. R. G. (2025). Lactoferrin as a candidate multifunctional therapeutic in synucleinopathies. Brain Sciences, 15(4), 380. https://doi.org/10.3390/brainsci15040380
- Bolesławska, I., Bolesławska-Król, N., Jakubowski, K., Przysławski, J., & Drzymała-Czyż, S. (2025). Lactoferrin — A regulator of iron homeostasis and its implications in cancer. Molecules, 30(7), 1507. https://doi.org/10.3390/molecules30071507
- Buccigrossi, V., de Marco, G., Bruzzese, E., Ombrato, L., Bracale, I., Polito, G., & Guarino, A. (2007). Lactoferrin induces concentration-dependent functional modulation of intestinal proliferation and differentiation. Pediatric Research, 61(4), 410–414. https://doi.org/10.1203/pdr.0b013e3180332c8d
- Colombo, J., Harris, C.L., Wampler, J.L., Zhuang, W., Shaddy, D.J., Liu, B.Y., & Wu, S.S. (2023). Improved neurodevelopmental outcomes at 5.5 years of age in children who received bovine milk fat globule membrane and lactoferrin in infant formula through 12 months: A randomized controlled trial. The Journal of Pediatrics, 261, 113483. https://doi.org/10.1016/j.jpeds.2023.113483
- Conesa, C., Bellés, A., Grasa, L., & Sánchez, L. (2023). The role of lactoferrin in intestinal health. Pharmaceutics, 15(6), 1569. https://doi.org/10.3390/pharmaceutics15061569
Donovan, S.M. (2016). The role of lactoferrin in gastrointestinal and immune development and function: A preclinical perspective. The Journal of Pediatrics, 173(Suppl.), S16–S28. https://doi.org/10.1016/j.jpeds.2016.02.072 - Elzoghby, A.O., Abdelmoneem, M.A., Hassanin, I.A., Abd Elwakil, M.M., Elnaggar, M.A., Mokhtar, S., Fang, J.-Y., & Elkhodairy, K.A. (2020). Lactoferrin, a multi-functional glycoprotein: Active therapeutic, drug nanocarrier & targeting ligand. Biomaterials, 263, 120355. https://doi.org/10.1016/j.biomaterials.2020.120355
- Gallo, V., & Antonini, G. (2024). Controversial role of lactoferrin in cancer: A narrative review. Biomedicine & Pharmacotherapy, 181, 117743. https://doi.org/10.1016/j.biopha.2024.117743
- Gruden, Š., & Poklar Ulrih, N. (2021). Diverse mechanisms of antimicrobial activities of lactoferrins, lactoferricins, and other lactoferrin-derived peptides. International Journal of Molecular Sciences, 22(20), 11264. https://doi.org/10.3390/ijms222011264
- He, J., & Furmanski, P. (1995). Sequence specificity and transcriptional activation in the binding of lactoferrin to DNA. Nature, 373(6516), 721–724. https://doi.org/10.1038/373721a0
- Hidalgo, I.J., Raub, T.J., & Borchardt, R.T. (1989). Characterization of the human colon carcinoma cell line (Caco-2) as a model system for intestinal epithelial permeability. Gastroenterology, 96(3), 736–749. https://doi.org/10.1016/S0016-5085(89)80072-1
- Huang, H.-C., Lin, H., & Huang, M.-C. (2019). Lactoferrin promotes hair growth in mice and increases dermal papilla cell proliferation through Erk/Akt and Wnt signaling pathways. Archives of Dermatological Research, 311(5), 411–420. https://doi.org/10.1007/s00403-019-01920-1
- Huang, N., Bethell, D., Card, C., Cornish, J., Marchbank, T., Wyatt, D., Mabery, K., & Playford, R. (2008). Bioactive recombinant human lactoferrin, derived from rice, stimulates mammalian cell growth. In Vitro Cellular & Developmental Biology – Animal, 44(10), 464–471. https://doi.org/10.1007/s11626-008-9136-7
- Jang, Y.-S., Park, S.-H., Kang, S.-G., Lee, J.-S., Ko, H.-J., & Kim, P.-H. (2023). Combined treatment with TGF-β1, retinoic acid, and lactoferrin robustly generate inducible Tregs (iTregs) against high affinity ligand. Immune Network, 23(5), e37. https://doi.org/10.4110/in.2023.23.e37
- Jiang, R., & Lönnerdal, B. (2014). Transcriptomic profiling of intestinal epithelial cells in response to human, bovine and commercial bovine lactoferrins. BioMetals, 27(5), 831–841. https://doi.org/10.1007/s10534-014-9746-3
- Jiang, R., Du, X., & Lönnerdal, B. (2014). Comparison of bioactivities of talactoferrin and lactoferrins from human and bovine milk. Journal of Pediatric Gastroenterology and Nutrition, 59(5), 642–652. https://doi.org/10.1097/MPG.0000000000000481
- Kopaeva, M. Y., Cherepov, A. B., Nesterenko, M. V., & Zarayskaya, I. Y. (2021). Pretreatment with human lactoferrin had a positive effect on the dynamics of mouse nigrostriatal system recovery after acute MPTP exposure. Biology, 10(1), 24. https://doi.org/10.3390/biology10010024
- Kosznik-Kwaśnicka K., Leszczyńska U., & Piechowicz L. (2026). Lactoferrin bridges antimicrobial and healing responses in Staphylococcus aureus skin infections. Frontiers in Microbiology. 16:1753483. https://doi.org/10.3389/fmicb.2025.1753483
- Kruzel, M.L., Actor, J.K., Zimecki, M., Wise, J., Płoszaj, P., Mirza, S., Kruzel, M., Hwang, S.-A., Ba, X., & Boldogh, I. (2013). Novel recombinant human lactoferrin: Differential activation of oxidative stress related gene expression. Journal of Biotechnology, 168(4), 666–675. https://doi.org/10.1016/j.jbiotec.2013.09.011
- Li, Y.-Q., & Guo, C. (2021). A review on lactoferrin and central nervous system diseases. Cells, 10(7), 1810. https://doi.org/10.3390/cells10071810
- Li, B., Zhang, B., Liu, X., Zheng, Y., Han, K., Liu, H., Wu, C., Li, J., Fan, S., Peng, W., Zhang, F., & Liu, X. (2021). The effect of lactoferrin in aging: Role and potential. Food & Function, 13(2), 501–513. https://doi.org/10.1039/D1FO02750F
- Liao, Y., Jiang, R., & Lönnerdal, B. (2012). Biochemical and molecular impacts of lactoferrin on small intestinal growth and development during early life. Biochemistry and Cell Biology, 90(3), 476–484. https://doi.org/10.1139/o11-075
- Lönnerdal, B., Du, X., & Jiang, R. (2020). Biological activities of commercial bovine lactoferrin sources. Biochemistry and Cell Biology, 99(1), 35–46. https://doi.org/10.1139/bcb-2020-0182
- Lönnerdal, B., Jiang, R., & Du, X. (2011). Bovine lactoferrin can be taken up by the human intestinal lactoferrin receptor and exert bioactivities. Journal of Pediatric Gastroenterology and Nutrition, 53(6), 606–614. https://doi.org/10.1097/MPG.0b013e318230a419
- MacManus, C.F., Collins, C.B., Nguyen, T.T., Alfano, R.W., Jedlicka, P., & de Zoeten, E.F. (2017). VEN-120, a recombinant human lactoferrin, promotes a regulatory T cell (Treg) phenotype and drives resolution of inflammation in distinct murine models of inflammatory bowel disease. Journal of Crohn’s and Colitis, 11(9), 1101–1112. https://doi.org/10.1093/ecco-jcc/jjx056
- Manzoni, P. (2016). Clinical benefits of lactoferrin for infants and children. The Journal of Pediatrics, 173(Suppl.), S43–S52. https://doi.org/10.1016/j.jpeds.2016.02.075
- Pan, S., Weng, H., Hu, G., Wang, S., Zhao, T., Yao, X., Liao, L., Zhu, X., & Ge, Y. (2021). Lactoferrin may inhibit the development of cancer via its immunostimulatory and immunomodulatory activities (Review). International Journal of Oncology, 59(5), 85. https://doi.org/10.3892/ijo.2021.5265
- Park, S.Y., Jeong, A.-J., Kim, G.-Y., Jo, A., Lee, J.E., Leem, S.-H., Yoon, J.-H., Ye, S.K., & Chung, J.W. (2017). Lactoferrin protects human mesenchymal stem cells from oxidative stress-induced senescence and apoptosis. Journal of Microbiology and Biotechnology, 27(11), 1877–1884. https://doi.org/10.4014/jmb.1707.07040
- Park, H.-W., Park, S.-H., Jo, H.-J., Kim, T.-G., Lee, J. H., Kang, S.-G., Jang, Y.-S., & Kim, P.-H. (2020). Lactoferrin induces tolerogenic bone marrow-derived dendritic cells. Immune Network, 20(5), e38. https://doi.org/10.4110/in.2020.20.e38
- Rai, D., Adelman, A.S., Zhuang, W., Rai, G.P., Boettcher, J., & Lönnerdal, B. (2014). Longitudinal changes in lactoferrin concentrations in human milk: A global systematic review. Critical Reviews in Food Science and Nutrition, 54(12), 1539–1547. https://doi.org/10.1080/10408398.2011.642422
- Rascón-Cruz, Q., Siqueiros-Cendón, T. S., Siañez-Estrada, L. I., Villaseñor-Rivera, C. M., Ángel-Lerma, L. E., Olivas-Espino, J. A., León-Flores, D. B., Espinoza-Sánchez, E. A., Arévalo-Gallegos, S., & Iglesias-Figueroa, B. F. (2025). Antioxidant potential of lactoferrin and its protective effect on health: An overview. International Journal of Molecular Sciences, 26(1), 125. https://doi.org/10.3390/ijms26010125
- Rizzi, M., Manzoni, P., Germano, C., Quevedo, M.F., & Sainaghi, P.P. (2025). Lactoferrin, a natural protein with multiple functions in health and disease. Nutrients, 17(21), 3403. https://doi.org/10.3390/nu17213403
- Schirmbeck, G. H., Sizonenko, S., & Sanches, E. F. (2022). Neuroprotective role of lactoferrin during early brain development and injury through lifespan. Nutrients, 14(14), 2923. https://doi.org/10.3390/nu14142923
- Shini, V.S., Udayarajan, C.T., & Nisha, P. (2022). A comprehensive review on lactoferrin: A natural multifunctional glycoprotein. Food & Function, 13(23), 11954–11972. https://doi.org/10.1039/D2FO02371G
- Siqueiros-Cendón, T., Arévalo-Gallegos, S., Iglesias-Figueroa, B. F., García-Montoya, I. A. (2014). Immunomodulatory effects of lactoferrin. Acta Pharmacologica Sinica, 35, 557–566. https://doi.org/10.1038/aps.2013.200
- Skoog, E.C., Castagna, V.F., Omer, S., Madigan, J., Flagg, V., Burrick, K., Jiang, R., Du, X., Lönnerdal, B., & Schnitzler, A. (2025). Structure and function of fermentation-derived bovine lactoferrin produced from Komagataella phaffii. Biochemistry and Cell Biology, 103, 1–17. https://doi.org/10.1139/bcb-2024-0105
- Suzuki, Y.A., Wong, H., Ashida, K.-Y., Schryvers, A.B., & Lönnerdal, B. (2008). The N1 domain of human lactoferrin is required for internalization by Caco-2 cells and targeting to the nucleus. Biochemistry, 47(41), 10915–10920. https://doi.org/10.1021/bi8012164
Takayama, Y., & Aoki, R. (2012). Roles of lactoferrin on skin wound healing. Biochemistry and Cell Biology, 90(3), 497–503. https://doi.org/10.1139/o11-054 - Takenaka, T., Harada, N., Kuze, J., Chiba, M., Iwao, T., & Matsunaga, T. (2014). Human small intestinal epithelial cells differentiated from adult intestinal stem cells as a novel system for predicting oral drug absorption in humans. Drug Metabolism and Disposition, 42(11), 1947–1954. https://doi.org/10.1124/dmd.114.059493
- Tang, L., Cui, T., Wu, J.J., Liu-Mares, W., Huang, N., & Li, J. (2010a). A rice-derived recombinant human lactoferrin stimulates fibroblast proliferation, migration, and sustains cell survival. Wound Repair and Regeneration, 18(1), 123–131. https://doi.org/10.1111/j.1524-475X.2009.00563.x
- Tang, L., Wu, J.J., Ma, Q., Cui, T., Andreopoulos, F.M., Gil, J., Valdes, J., Davis, S.C., & Li, J. (2010b). Human lactoferrin stimulates skin keratinocyte function and wound re-epithelialization. British Journal of Dermatology, 163(1), 38–47. https://doi.org/10.1111/j.1365-2133.2010.09748.x
- Vishwanath-Deutsch, R., Dallas, D.C., Besada-Lombana, P., Katz, L., Conze, D., Kruger, C., Clark, A.J., Peterson, R., & Malinczak, C.-A. (2024). A review of the safety evidence on recombinant human lactoferrin for use as a food ingredient. Food and Chemical Toxicology, 189, 114727. https://doi.org/10.1016/j.fct.2024.114727
- Wang, B., Timilsena, Y.P., Blanch, E., & Adhikari, B. (2019). Lactoferrin: Structure, function, denaturation and digestion. Critical Reviews in Food Science and Nutrition, 59(4), 580–596. https://doi.org/10.1080/10408398.2017.1381583
- Wolf, J.S., Li, G., Varadhachary, A., Petrak, K., Schneyer, M., Li, D., Ongkasuwan, J., Zhang, X., Taylor, R.J., Strome, S.E., & O’Malley, B.W. (2007). Oral lactoferrin results in T cell–dependent tumor inhibition of head and neck squamous cell carcinoma in vivo. Clinical Cancer Research, 13(5), 1601–1610. https://doi.org/10.1158/1078-0432.CCR-06-2008
- Xie, T., Qiao, W., Jia, T., & Kaku, K. (2024). Skin care function of lactoferrin was characterized using recombinant human epidermal model. Cosmetics, 11(3), 98. https://doi.org/10.3390/cosmetics11030098
- Xu, S.-F., Zhang, Y.-H., Wang, S., Pang, Z.-Q., Fan, Y.-G., Li, J.-Y., Wang, Z.-Y., & Guo, C. (2018). Lactoferrin ameliorates dopaminergic neurodegeneration and motor deficits in MPTP-treated mice. Redox Biology, 21, 101090. https://doi.org/10.1016/j.redox.2018.101090
- Xu, S.-F., Cui, J.-H., Liu, X., Pang, Z.-Q., Bai, C.-Y., Jiang, C., Luan, C., Li, Y.-P., Zhao, Y., You, Y.-M., & Guo, C. (2024). Astrocytic lactoferrin deficiency augments MPTP-induced dopaminergic neuron loss by disturbing glutamate/calcium and ER-mitochondria signaling. Free Radical Biology and Medicine. https://doi.org/10.1016/j.freeradbiomed.2024.10.284
- Yagi, M., Suzuki, N., Takayama, T., Arisue, M., Kodama, T., Yoda, Y., Otsuka, K., & Ito, K. (2009). Effects of lactoferrin on the differentiation of pluripotent mesenchymal cells. Cell Biology International, 33(3), 283–289. https://doi.org/10.1016/j.cellbi.2008.11.013
- Zhang, Z., Lu, M., Chen, C., Tong, X., Li, Y., Yang, K., Lv, H., Xu, J., & Qin, L. (2021). Holo-lactoferrin: The link between ferroptosis and radiotherapy in triple-negative breast cancer. Theranostics, 11(7), 3167–3182. https://doi.org/10.7150/thno.52028
- Zlatina, K., & Galuska, S. P. (2021). The N-glycans of lactoferrin: More than just a sweet decoration. Biochemistry and Cell Biology, 99(1), 117–127. https://doi.org/10.1139/bcb-2020-0106